Calcium (Ca) is an essential macro-mineral involved in many physiological processes including skeletal and exoskeleton mineralization, muscle contraction, nerve transmission, osmoregulation, and coagulation. While a lack of Ca can have harmful effects on the performance of fish and shrimp, supplementation in the feed is, for most of the time, not necessary, as the latter can be absorbed directly from the water. On the contrary, an excess of Ca in the feed can affect the absorption of other nutrients and minerals including phosphorus (P).
The Functions of Calcium
Bone mineralization
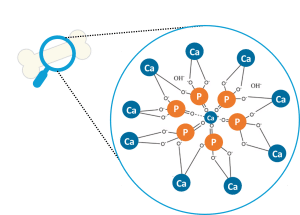
As in terrestrial animals, Ca is the most present macro-mineral in fish body, with levels ranging from 1.5 to 4% of dry weight and 0.05 to 0.2% of fresh weight depending on the species and development stage (Lall et al., 2002; NRC, 2011). Ca is mainly stored in bones such as terrestrial vertebrates, but also in scales, which are rich in Ca. They can serve as a stock of Ca that can be mobilized during periods of increased Ca needs, such as sexual maturation or fasting periods (Persson et al., 1998). Tilapia and Sturgeon have thick, highly calcified scales, thus increasing Ca levels to about 3.5% of dry weight in Nile tilapia (Oreochromis niloticus) compared to 2.5% in Atlantic salmon (Salmo salar). In bones, Ca is, together with P, first deposited as tricalcium phosphate (Ca3(PO4)2) before crystallization into hydroxyapatite (Ca10(PO4)6(OH)2) and integration into the organic matrix (Figure 1). As an essential component of bone along with P, Ca plays a crucial role in bone formation and maintenance. However, no studies clearly show the importance of Ca in bone formation in fish in contrast to P deficiency (Drábiková et al., 2021), because it is difficult to induce Ca deficiency, due to Ca absorption from water.
Figure 1. Hydroxyapatite crystals
Exoskeleton and moulting
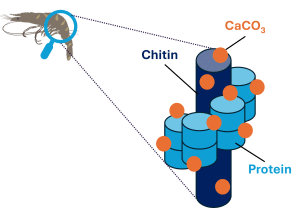
Arthropods such as shrimp do not have an internal skeleton like vertebrates but an external skeleton called exoskeleton or cuticle. As it grows, the shrimp must get rid of this rigid exoskeleton that cannot expand and prevents shrimp growth, before forming a new one. This cyclical physiological process called moulting or ecdysis, is dependent on the availability of Ca. Indeed, the exoskeleton of arthropods is made up of an organic matrix of chitin (20 to 30% of the dry weight) and proteins as well as minerals including Mg and especially calcium carbonate (CaCO3) which represents 20 to 30% of the dry weight of the exoskeleton (Brannon et al., 1979; Stevenson et al., 1985) (Figure 2). CaCO3 is deposited on the organic matrix and contributes to its thickening and resistance. During moulting, part of the Ca of the old exoskeleton is mobilized and stored in the hemolymph and hepatopancreas, reducing its rigidity and facilitating exuviation and removal. Then, the new cuticle, which is then flexible and vulnerable, rapidly absorbs Ca to allow calcification and harden it (Stevenson et al., 1985). It is estimated that about 38% of the nutrients in the old cuticle are recycled (Lemos et al., 2021). In addition, many species eat the ancient cuticle, however the digestibility of its minerals is unknown (Zanotto & Wheatly, 2002). Shrimp moulting is critical in protecting shrimp from predators, pathogens, and injury. Thus, a Ca deficiency can affect shrimp moulting, impacting their growth performance and vulnerability.
Figure 2. Chitin-protein fibers
Intake and Ca requirements of aquaculture species
Calcium absorption in fish and shrimp
Ca is a mineral that is highly present in seawater and to a lesser extent in freshwater. Ca can be absorbed from the gastrointestinal tract as well as from the gills (Flik & Verbost, 1993). It also appears that Ca may be absorbed directly through the skin (Marshall et al. 1992). However, the gills remain the main organ of Ca absorption in fish (Flik et al., 1995), since the absorption surface is 10 to 60 times larger for the gills than for the skin (Parry, 1966). Although the gastrointestinal tract has a less important role in the uptake of Ca, it can still contribute significantly during periods of increased Ca requirements (gonad maturation, egg formation) and rapid growth (Flik et al., 1993) or in water lacking Ca (Robinson et al., 1986). In addition, other water parameters impact the absorption of Ca by the gills. Indeed, the absorption of Ca from water is dependent on salinity, as observed in larvae of Gilthead bream (Sparus aurata) which have seen their absorption of Ca increases with salinity (Guerreiro et al., 2004).
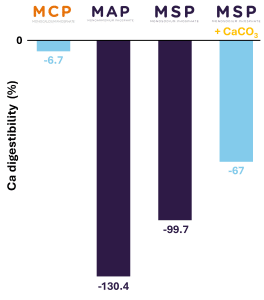
In the Whiteleg shrimp (Litopenaeus vannamei), Phosphea was able to observe similar results in a study in collaboration with the University of Sao Paulo (Coelho et al., 2024). Indeed, the digestibility values of Ca obtained were negative, demonstrating that Ca is mainly absorbed from water and not from the feed (Figure 3).
The use of calcium phosphates such as monocalcium phosphate (MCP) may only reduce the uptake of Ca from water compared to non-calcium phosphates such as monosodium phosphate (MSP), monoammonium phosphate (MAP) or magnesium phosphate (MgP) (Coelho et al., 2024).
Figure 3. Calcium digestibility in Whiteleg shrimp
Dietary calcium requirements
Since fish can absorb Ca from water, dietary Ca intake is usually not necessary. Studies show that freshwater fish fed Ca-deficient diets grow normally if the water contains enough Ca (Hwang et al., 1996). However, if the water is deficient in Ca, fish growth may be stunted (Rodgers 1984). Nevertheless, it has been observed that dietary Ca may be essential for some freshwater species such as grass carp (Ctenopharyngodon idella), silver catfish (Rhamdia quelen) or Nile tilapia. In the latter, as well as in brook trout (Salvelinus fontinalis), it has been observed that the absorption of dietary Ca varies according to the Ca concentrations of the water. The richer the water in Ca, the less dietary Ca contributes to satisfying the needs of fish (Phillips, 1959; Vonck et al., 1998).
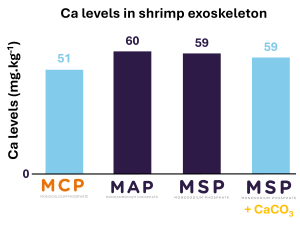
Since seawater contains more Ca, it is considered that marine species have no need for dietary Ca. However, there are studies demonstrating that dietary Ca supplementation is necessary for certain species such as the tiger puffer fish (Takifugu rubripes) (Hossain & Furuichi, 1998). As with fish, Whiteleg shrimp and Giant tiger shrimp (Penaeus monodon) do not have any particular dietary requirements (Peñaflorida et al., 1999; Davis et al., 1993). In Phosphea study (Coelho et al., 2024), the analysis of the composition of the exoskeleton shows that the use of non-calcium phosphates has no effect on the mineralization of Ca in the cuticle compared to MCP. Showing that non-calcium phosphate such as MSP or MAP can be used without deleterious effect on Ca levels in shrimp exoskeleton (Figure 4).
Figure 4. Ca levels in shrimp exoskeleton
Effects of excess calcium
On the contrary, Ca intake can even have deleterious effects on the growth of shrimp and fish. Indeed, as in land-based livestock, Ca affects the digestibility of other nutrients, including other minerals, such as P, Magnesium (Mg) and Zinc (Zn). Dietary Ca decreases the absorption of these minerals, probably by competing with other cations for absorption (Song et al., 2016) and binding to P, forming complexes that are not absorbable by monogastric animals (Andrews et al., 1973; Cowey & Sargent, 1979). In fish, dietary Ca has been shown to decrease the absorption of P. Nakamura has observed a negative linear correlation between P absorption and the Ca content of the feed in carp (Nakamura, 1982). This negative effect of dietary Ca, affecting growth and feed efficiency, has also been observed in other species including Rainbow trout (Porn-Ngam et al., 1993), Japanese eel (Nose and Arai, 1979), Channel catfish (Gatlin & Phillips, 1989), and King salmon (Richardson et al., 1985).
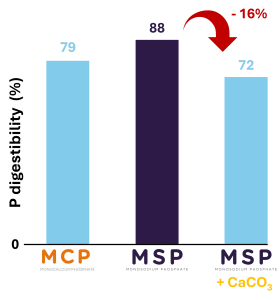
In Whiteleg shrimp, Phosphea study clearly shows the negative effect of dietary Ca on the digestibility of P (Coelho et al., 2024). Indeed, the addition of Ca carbonate (CaCO3) to shrimp feed containing monosodium phosphate (MSP), which is a highly digestible inorganic feed phosphate in shrimp, significantly reduces the digestibility of P (Figure 5). The addition of CaCO3 to the feed decreases the digestibility of MSP P by more than 16%, from more than 88% to less than 72%.
Figure 5. Impact of Calcium on Phosphorus digestibility in Whiteleg shrimp
According to numerous studies, it seems essential to limit the intake of Ca in the feed to maintain a Ca/P ratio depending on the species and the stage of development to avoid the antinutritional effects of Ca. The Ca/P ratios by species are available in Table 1.
Table 1: Ca/P ratio requirements for aquaculture species

Conclusion
Based on the body of scientific evidence available, dietary Ca intake is not required for most freshwater and marine aquaculture species. However, some species may require additional Ca, including species with significant scale calcification such as tilapia or pufferfish.
- Since dietary Ca impacts the absorption of other nutrients, including P, it is preferable to limit its intake in the feed. It is therefore recommended to use sources of P free of Ca such as MSP or MAP which have a higher water solubility and digestibility than calcium phosphates such as DCP and MCP as well as organic sources such as bone meal or fishmeal which contain P and Ca in the form of tricalcium phosphate, not digestible for fish and shrimp.
- Since Ca deficiencies depend mainly on Ca concentrations in the water, it is preferable to directly increase Ca levels in the water by using a soluble Ca source such as CalseaPowder Advance (CPA), which provides assimilable Ca while avoiding the anti-nutritional effects of dietary Ca. In addition, a supplementation of Ca increases the hardness of the water and stabilizes the pH by acting as a buffer, improving water quality for fish and shrimp.
Author : Guillaume Morin